RESEARCH HIGHLIGHTS OF LAST 10 YEARS
1. Long-term lipid diet can generate non-colony forming persisters:
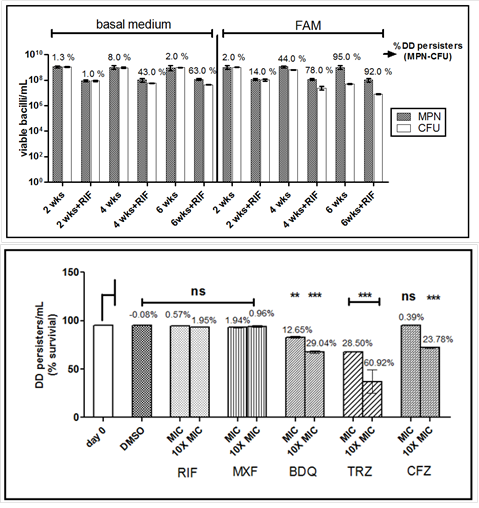
Persisters of Mycobacterium tuberculosis (Mtb) that fail to form colonies on agar media when de-stressed are termed as differentially detectable (DD) persisters. Since in the host, Mtb primarily survives by utilizing lipids, we used a long-term lipid diet model to induce DD persisters of M. tuberculosis. Persisters were induced by replacing the dextrose-containing medium with one containing fatty acids instead of dextrose (FAM). After 2, 4 or 6 weeks, CFU and most probable number assays were performed; the difference between the two gave an estimate of DD persisters. Since rifampicin has been shown to induce formation of DD persisters in vitro, one set of FAM cultures were also given short- term rifampicin stress after 2, 4 or 6 weeks. Fraction of DD persisters increased with time and rifampicin treatment enhanced the effect of fatty acids, at 2 and 4 weeks. At six weeks, even in the absence of rifampicin, ~95 % population were DD persisters. The DD persisters were vulnerable to drugs interfering with bacterial respiration such as thioridazine, bedaquiline and clofazimine. The study indicates potential formation of DD persisters of Mtb in a lipid-rich microenvironment in the host even before antibiotic therapy.
(Khan SR et al., Tuberculosis (Edinb). 2019).
2. Utilization of triacylglycerols during persistence:
During its persistence in the infected host, Mtb accumulates host-derived fatty acids in intra-cytoplasmic lipid inclusions as triacyl glycerols (TAG) which serve primarily as carbon and energy reserves. The Mtb genome codes for more than fifteen triacyl glycerol synthases (TGS), twenty four lipase/esterases and seven cutinase-like proteins. Hence we looked at the expression of the corresponding genes in intra-cellular bacilli persisting amidst of host TAG. We used the Mtb infected murine adipocyte model to ensure persistence and transcripts were quantified by real time reverse transcriptase PCR. Dormancy and glyoxylate metabolism was confirmed by the up-regulated expression of dosR and icl respectively by intra-adipocyte bacilli compared to in vitro growing bacilli. The study revealed that tgs1, tgs2, Rv3371 and mycolytransferase Ag85A are the predominant TGS, while lipF, lipH, lipJ, lipK, lipN, lipV, lipX, lipY, culp5, culp7 and culp6 are the predominant lipases/esterases used by Mtb for the storage and degradation of host derived fat. Moreover, it was observed that many of these enzymes are used by Mtb during active replication rather than during non-replicating persistence, indicating their probable function in cell wall synthesis.
(Rastogi S et al., Int J Mycobacteriol. 2016).

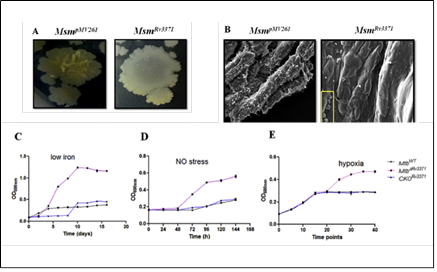
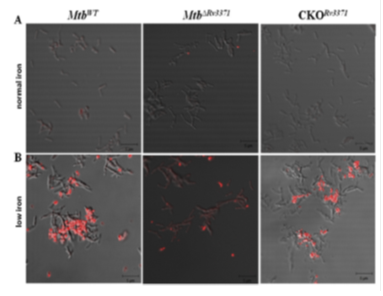
We identified that one of the TAG synthases of Mtb, Rv3371, which is specific to pathogenic mycobacteria, when expressed in M. smegmatis leads to modifications in colony morphotype, bacterial architecture, cell surface properties and elevated TAG levels. Rv3371 was found to largely localize in the cell membrane. The Rv3371 promoter is minimally active during exponential growth in vitro, however, is up-regulated under stationary phase, hypoxia, nutrient starvation, nitrosative stress, low iron, in IFN-γ activated macrophages and infected mice. The low iron-induced expression of Rv3371 is likely due to the de-repression by Rv1404, which is probably activated by ideR. An Rv3371 deletion mutant of Mtb showed impaired non-replicating persistence in vitro and altered sensitivity to anti-mycobacterial drugs. In low iron medium, the Rv3371 deletion mutant showed reduced formation of TAG containing extracellular vesicles. Therefore Rv3371 is likely involved in Mtb growth arrest and cell wall alterations during persistence.(Rastogi S et al., Tuberculosis (Edinb). 2017).
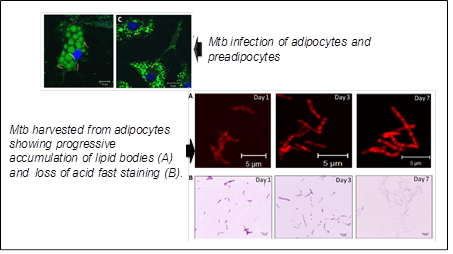
3. M. tuberculosis and adipocytes:
The adipocytes are one of the non-professional phagocytes postulated to be a haven for Mtb during persistence in the human host. We investigated Mtb infection of adipocytes in course of infection of mouse model. Mtb bacilli could be isolated form visceral, subcutaneous, peri- renal and mesenteric adipose depots of immunocompetent mice infected with Mtb intravenously. The bacilli could be isolated from adipocytes and the stromal vascular fraction, even though the numbers were significantly higher in the latter. The bacterial burden in the adipose depots was comparable to those in lungs in the early phase of infection. But with time, the burden in the adipose depots was either decreased or kept under control, despite the increasing burden in the lungs. Infected mice treated with standard anti tubercular drugs, despite effective elimination of bacterial loads in the lungs, continued to harbour Mtb in adipose depots at loads similar to untreated mice in the late infection phase. Mtb also induced changes in the immune cell constitution of the adipose tissue. The macrophage-specific marker F4/80 was detected in 6.7% of the total stromal vascular cells from infected mice compared to 0.4% in uninfected mice. Mature T-lymphocytes (CD3e+) contributed to 16% of the stromal vascular fraction from uninfected mice. Of these, 7.4% were CD4+ T-cells. However, in the adipose tissue of mice infected for two weeks, the T-cell numbers were reduced by half compared to uninfected mice and almost all were CD3e + CD4+ T cells. We have also shown Mtb can disseminate from lungs to distant adipose depots and vice versa.
.(Agarwal P et al., Microbes Infect. 2014 ; Agarwal P et al., Microb Pathog. 2016).
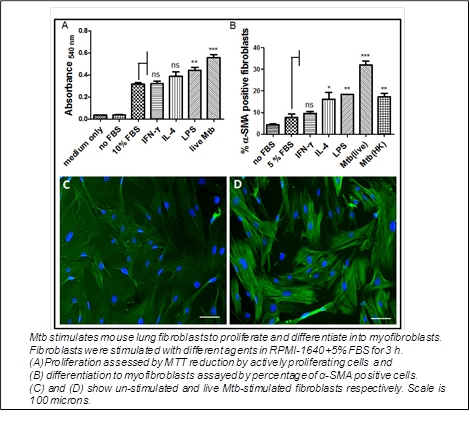
4. M. tuberculosis and lung fibroblasts:
Going with the classical role in wound healing, fibroblasts of the lung take part in the resolution of tubercular granulomas. They are totally absent in nascent granulomas, but surround necrotizing granulomas, and are the majority in healed granulomas. Lung fibroblasts may get infected with Mtb. Evidences suggested an immunomodulatory effect of fibroblasts on infected macrophages. Hence we looked at the role of primary mouse lung fibroblasts on naive or activated mouse bone marrow macrophages infected with Mtb and the effect of infection on fibroblast properties. We observed that with fibroblasts in the vicinity, infected naive macrophages restricted the bacterial growth, while activated macrophages turned more bactericidal with concomitant increase in nitrite production. Neutralizing IL-1α in fibroblast supernatant reduced the nitrite production by infected macrophages. Secretion of IL-6 and MCP-1 was down- regulated, while TNF-α was up-regulated in infected naive macrophages. In infected activated macrophages, the secretion of IL-6 was up-regulated, while that of MCP-1 and TNF-α was unaffected. The ‘fibroblast effects’ were enhanced when the fibroblasts too were infected. Mtb induced IL-1 secretion and pro-fibrotic responses by fibroblasts. Mtb-induced myofibroblast conversion was blocked by rapamycin suggesting the involvement of signalling via mTOR.
(Verma SC et al., Tuberculosis (Edinb). 2016).